ТЕЛЕСКОП “ДЖЕЙМС УЭББ” И ПРАВОСЛАВНОЕ БОГОСЛОВИЕ
II часть
Валентин Велчев
Несколько слов о Боге
Согласно Библии, Бог есть Дух. Его естество (природа,
субстанция) духовно, то есть совершенно отлично от материи. Из того, что Бог
нематериален, следует, что никакими средствами науки и техники невозможно
установить Его присутствие (так, как с их помощью можно зарегистрировать
наличие какого-либо физического поля, например). Другими словами, Его
существование не может быть доказано прямо – только косвенно можно судить о
том, что Он действительно существует.
Бог трансцендентен,
то есть находится вне материального пространственно-временного континуума, а
также имманентен (т.е. вездесущ). Это значит, что Он присутствует всегда
и везде, но оставаясь отдельным и независимым от всего. Он не безличная сила
или энергия, а личностный Бог с разумом, чувством, волей и ярко выраженным
характером, Который не только мудро создал, но и перманентно правит всем
мировым порядком. В конечном счете Бог держит все сущее под Своим полным
контролем и обладает суверенным правом управлять миром различными способами. В
какой-то момент Он может оставить события развиваться в согласии с
естественными законами, а в другой – сотворить чудо, чтобы помочь людям или с
какой-то другой целью.
Библия
представляет Бога как три единосущные Лица: Отец, Сын и Дух Святой; именно в
силу этого Бог является Единством в Троице. Эта истина, открытая нам в Писании,
является тайной сверхразумной и не имеющей параллелей в человеческом опыте.
Поэтому интеллект так же неспособен проникнуть в нее, как и язык – передать ее
доступным образом. Наша дилемма – как выразить невыразимое, чтобы обосновать
свою веру – отражена в словах Августина Блаженного: “Три Лица – не потому, что
можно говорить об этом, но потому, что нельзя оставаться в молчании”.
Бог существует бесконечно (не забудем о том, что Он вне времени), при этом
никогда не было причины, породившей Его. Другими словами, Бог существует вечно
Сам по Себе. Таким образом вопрос “Кто создал Бога?”, постоянно задаваемый
напоследок некоторыми учеными, совершенно несостоятелен.
Творение мира
Согласно христианскому учению, Триединый Бог в начале
вызвал к бытию все сущее – материальное и духовное, из ничего (лат. ex nihilo).
Хотя слова “из ничего” не встречаются в Библии, сама идея об этом наличествует
во многих местах Ветхого и Нового Заветов[1]. В
богословии выражение “из ничего” используется, чтобы пояснить, что Бог не
использовал какую бы то ни было первоначальную субстанцию. Необходимо уточнить,
что “ничто” в принципе не имеет бытия, что означает, что перед “началом”
существовал только Бог. Разумеется, после сотворения материи в дальнейшем Бог
образовал растения, животных и людей из ее компонентов, то есть из атомов
химических элементов, наличествовавших в морях и на суше.
Творение есть акт Его суверенной воли, а не действие,
продиктованное какими-либо внешними соображениями или необходимостью. Для того,
чтобы призвать мир к бытию, Он использует только Свое Слово. В первой главе
Книги Бытия мы читаем, что Бог говорит и слова Его мгновенно превращаются в
действительность. Ему достаточно сказать “Да будет свет!” – и свет моментально
является и освещает небесные просторы. Согласно бл. Августину и св. Максиму
Исповеднику, “логосы” (идеи) в Божьем уме являются “архетипами” (прообразами)
каждой частицы Вселенной. Они объективированы именно как “тропосы”, выражающие
творческий замысел в природе вещей. Поэтому свет, например, имеет
корпускулярно-волновой характер, поляризуется, движется с определенной
скоростью и т.д.
Возможна ли верификация библейского мировоззрения?
Ричард Докинз
заявляет: “Естественный отбор, слепой,
несознательный, автоматический процесс, который был открыт Дарвином и о котором
мы сейчас знаем, что он объясняет существование и мнимо целесообразную форму
всего живого, не следует никакому намерению. В нем нет разума и разумного ока.
Он не планирует будущее. Он не смотрит и не предвидит, он вообще не зряч. Если
можно сказать, что он играет роль часовщика в природе, то это слепой
часовщик”[2]. А
американский когнитивный философ Дэниел Деннетт указывает на естественный отбор
как на фактор, объясняющий совершенство всех природных структур (физических,
химических, биологических) и таким образом ставит его в основу всей
натуралистической парадигмы: “Теория
естественного отбора показывает, как любая особенность естественного мира может
быть продуктом слабого, несправедливого, нетелеологического и в конечном
счете механического процесса дифференциальной репродукции в течение долгих
периодов времени”[3].
Мы совершенно согласны с
утверждением Докинза, что нужно найти способ эмпирической проверки эволюционной
и библейской космологических моделей происхождения Вселенной (см. I часть этой статьи). Более того, поскольку Докинз – воинствующий атеист, то
вряд ли может возникнуть подозрение, что подобный критерий придуман им для
того, чтобы поддержать христианство. Далее мы предложим подход, позволяющий во
второй части статьи проверить также и библейскую модель, но, поскольку проблема
достаточно сложна (но, к счастью, не неразрешима!), необходимо предварительно
уточнить некоторые общие положения.
Библейские авторы и ученые говорят
на разных языках. Язык науки строго
специализирован и техничен. Он развивался, чтобы служить совершенно конкретным
целям, и нередко его могут понять только специалисты. Язык Библии
феноменологичен – он показывает вещи такими, какими они выглядят. Другими
словами, язык Библии по отношению к природе популярен, это язык, на котором
обыкновенные люди говорили в соответствующую эпоху и в соответствующее время.
Первые главы Книги Бытия не защищают и не опровергают теории Аристотеля,
Птолемея, Коперника, Ньютона или Эйнштейна. Библия в принципе отказывается
говорить о подобных теориях в какой бы то ни было области – астрономии,
геологии, физике, химии, биологии. В Библии отсутствуют научные постулаты. Она
просто описывает природу, не объясняя ее в научном аспекте. Вот почему, говоря
о “библейской космологической модели”, мы будем
иметь в виду основные черты того, как Моисей описал сотворение Земли, Солнца,
Луны и звезд, а также принципиальную разницу между сознательной деятельностью и
слепым случаем.
Как правильно советуют некоторые богословы,
человек не должен умозрительно пытаться понять, как возникла Вселенная,
потому что Бог в состоянии создать окружающий нас мир способом, абсолютно
недоступным нашему разуму и воображению, т.е. мы не должны накладывать
ограничения на всемогущество Божие. Как говорит известный православный богослов
Иустин Попович: “Способ, которым
создан мир, настолько сложен и таинственен в своей основе, что недоступен для
человеческого ума”. Ст. Хокинг и Л. Млодинов в своей книге “Высший замысел”, ссылаясь
на Ричарда Фейнмана, заявляют, что Вселенная могла появиться всеми возможными
способами.[4] Дополним, что божественный творческий акт
предполагает, что она возникла не только всеми возможными, но и “невозможными”
способами. Все же мы надеемся, что научный метод, раскрывающий перед нами одно
за другим чудеса природы, в скором времени раскроет нам и тайну Творения.
Основатель эмпирического подхода и
отец всей современной науки Галилео Галилей утверждает: “Главная цель Библии – поклонение Богу и спасение душ. [...] Но сотни
мест в ней учат нас тому, что слава и величие всемогущего Бога совершенно
видимы во всех Его творениях и могут быть прочитаны в открытой Книге природы”[5].
И действительно, с помощью современных научных средств – ускорителей
элементарных частиц, телескопов, наблюдающих сверхглубокий космос, детекторов
гравитационных волн, компьютерных симуляций и т.д. – мы в состоянии заглянуть
уже в первые мгновения мироздания. Поэтому мы попытаемся найти ответ на вопрос,
заданный Докинзом: “Является ли наша Вселенная делом разумного Творца, или
же она возникла и развилась чисто случайным образом? (см. I часть)”. Можно разграничить эти две точки зрения следующим образом:
А) Дарвин будет прав, если окажется,
что материя эволюционирует на основе множества вариаций, порожденных мутациями,
а естественный отбор отбирает наиболее удачные формы и таким образом приводит к
совершенствованию физических, химических и биологических структур. Ясно, что направление природных процессов в
данном случае – от хаоса к порядку.
Б) Премудрый
Бог должен был создать прекрасно устроенный и гармонический мир, в котором все просчитано
и с самого начала идет по строго определенному плану. Дополнительным
усложнением, однако, является то обстоятельство, что после грехопадения, по
выражению апостола Павла, “тварь
покорилась рабству тления (т.е.
разрушения)” (ср. Рим. 8:20-21), что
предполагает, что в мире будут наблюдаться также и процессы дезинтеграции (взрывающиеся
звезды, сталкивающиеся галактики и пр.). Но
направление процессов будет здесь совершенно противоположным – от порядка к
хаосу.
Библейская космологическая модель[6]
Рассказ Книги Бытия возвещает, что сотворение Земли и небесных светил произошло в первый и четвертый творческий день:
“В начале
сотворил Бог небо и землю. Земля же была безвидна и пуста, и тьма
над бездною, и Дух Божий носился над водою. И сказал Бог: да будет свет.
И стал свет. И увидел Бог свет, что он хорош, и отделил Бог свет от тьмы. И
назвал Бог свет днем, а тьму ночью. И был вечер, и было утро: день один. […]
И сказал
Бог: да будут светила на тверди небесной [для освещения земли и] для отделения
дня от ночи, и для знамений, и времен, и дней, и годов; и да будут они
светильниками на тверди небесной, чтобы светить на землю. И стало так. И создал
Бог два светила великие: светило большее, для управления днем, и светило
меньшее, для управления ночью, и звезды; и поставил их Бог на тверди
небесной, чтобы светить на землю, и управлять днем и ночью, и отделять свет от
тьмы. И увидел Бог, что это хорошо. И был вечер, и было утро: день четвертый”
(Быт. 1:1-14).
Почти все святые отцы считают, что “небо” в первом стихе обозначает невидимый мир – местопребывание престола Божия, окруженного ангелами, херувимами, серафимами и другими силами небесными. Этот трансцендентный мир более подробно описан в книгах Исаии, Иезекииля,
в Откровении и т.д., но мы не станем останавливаться на нем. В
отношении же слова “земля” мнения толкователей Библии разделяются примерно в двух направлениях:
А) Одни
считают, что, кроме нашей Земли, это слово
обозначает также и всю материю
в космосе, например в виде газо-пылевых облаков (тоже самое относится и к темной материи и энергии). В таком случае, по велению Божию в дальнейшем в них должны
были сформироваться небесные тела, движущиеся по своим орбитам и образующие планетные, звездные и
галактические системы. Все же, хотя и образованные во времени, массы и
орбитальные характеристики тел и систем должны были быть прекрасно просчитаны,
чтобы не было хаотических столкновений между ними и катастроф, что позволяет
ясно отграничить эту точку зрения от натуралистических моделей.
Б) Другие полагают, что слово “земля”
относится лишь к нашей планете, а Солнце, Луна и звезды появились в четвертый день, и, следовательно, Вселенная возникла сразу уже полностью упорядоченной.
В библейской модели “Большим
взрывом” можно считать
мгновенное сотворение пространственно-временного материального континуума, но,
в отличие от Стандартного сценария, здесь
все не стартует из одной
точки. Математическая попытка объединения общей теории
относительности и квантовой механики убедительно показала, что Вселенная не могла стартовать из точки нулевого размера с бесконечной
плотностью, что ведет к вопросу: “Каким был ее
первоначальный объем?” Как подчеркивает бразильский физик Джулиано Невис, “не существует никаких доказательств существования первичной сингулярности”[7], что
говорит о том, что космологи неправомерно экстраполируют события обратно во
времени до некоей исходной точки
Здесь можно сделать одно допущение, а именно: возможно, что космический
микроволновый фон (КМФ) является остатком того “света”, который
озарил небесные просторы в первый день Творения. Если это действительно так, то
мы могли бы еще раз уточнить,
какой из двух библейских сценариев вероятнее всего был реализован на практике.
Согласно
первому варианту, Земля и газо-пылевые облака появились до КМФ, а согласно
второму – вся материя в виде готовых звездных систем появилась после него.
Кроме того, при постепенном образовании тел и формировании космических структур
характер гравитационных волн и флуктуаций (а, может быть, и поляризации) КМФ
должен отличаться от того, который бы наблюдался при мгновенном возникновения Вселенной.
Но чтобы
найти ответ на эту загадку, придется подождать еще лет десять, до момента,
когда НАСА и ESA планируют вывести на орбиту самый совершенный детектор
гравитационных волн – LISA.[8] Как выражаются
некоторые ученые, „он будет в состоянии снять отпечатки пальцев Бога, оставшиеся
на ткани космоса еще в первые моменты Сотворения”. И, действительно, детектор
LISA будет настолько высокоточным, что сможет уловить ударные волны еще первой
трилионной части секунды после Большого взрыва. (Для сравнения, измеренное
предыдущим аппаратом WMAP космическое фоновое излучение восходит к моменту,
когда исполнилось 380 тысяч лет от начала, – это время, когда, как считают,
начали рекомбинироваться атомы.)
Научная проблема: с точки зрения теории
космические системы необъяснимы!!
Как мы
заявили еще в самом начале, предметом этой статьи является вопрос: возможно ли
вообще, чтобы случайным образом получился этот исключително красивый и сложный иерархический
порядок небесных образований – планетарных, звездных, галактических и пр. – как
и его огромная устойчивость во времени? Дело в том, что даже если налицо только
три небесные тела со случайно выбранными массами и орбитальными характеристиками,
то их система оказывается неустойчивой, а что говорить о системах из большего
количества тел.
В конце XVII века Исаак Ньютон открыл закон всемирного притяжения и
сумел объяснить им движение планет вокруг Солнца. В начале он решил задачу двух
тел – например, какой-нибудь планеты и Солнца (без влияния других тел), и таким
образом показал, что орбита такой планеты, как Земля, будет образовывать
определенный эллипс, который может быть точно описан математически. Но когда к
этой системе подключается и Луна, то оказывается, что нельзя получить общее
математическое решение о положении трех тел, движущихся в пространстве
единственно под влиянием своего взаимного гравитационного притяжения. Немного
позже Анри Пуанкаре доказал, что подобные взаимодействия хаотичны (и тем самым
положил начало „теории хаоса“[9]), в смысле, что
конечный результат по существу случаен и непредсказуем, так что эволюция системы
из трех (и большего числа) тел невозможно предвидеть в каких бы то ни было
временных масштабах.
Компьютерные симуляции показали, что
взаимодействие в системе из трех тел протекает в двух фазах: в начале идет хаотическая
фаза, во время которой все три тела притягиваются сильно друг к другу, пока
одно из них не будет отброшено далеко от других, которые после этого займут
эллиптические орбиты вокруг общего центра масс. Если третье тело останется на привязанной
орбите, то оно в конечном счете возвратится обратно в двойную систему, после
чего первая фаза наступит снова. Этот тройной танец может закончиться тогда,
когда во второй фазе одно из тел удалится по непривязанной, т.е. гиперболической
орбите и начнет удаляться бесконечно в пространство, чтобы не возвратиться
никогда. Есть известная вероятность того, что два из трех тел могут слиться
после столкновения, но во всех случаях система будет неустойчивой во времени.[10]
То же в силе и для любой системы из N тел, причем в этом случае выброшенное третье тело скорее всего будет
захвачено гравитационным полем какого-нибудь из других тел, в результате чего
начнутся непрерывные столкновения и слияния тел. В конечном счете все вещество
системы соберется в одно массивное тело, вместившее в себя (почти) все остальные
(см. третью гиперссылку в прим. [11] I части). Образование сбалансированной системы возможно лишь в пренебрежимо
ничтожном проценте случаев, причем при увеличении числа тел этот процент
уменьшается экспоненциально, что означает, что на практике осуществление такого
варианта ниже любой допустимой вероятности!
Над разрешением
вопроса, стабильна ли Солнечная система, в прошлом особенно активно работали
такие выдающиеся математики и астрономы как Лаплас, Лагранж, Пуанкаре и др. И
все же только к середине XX века стало возможным разработать математические
методы и средства, на основе которых советские ученые А. Колмогоров, В. Арнольд
и Ю. Мозер дали доказательство стабильности Солнечной системы. До недавнего
времени считалось, что наша планетная система может сохраняться около триллиона
лет, но недавняя работа (2020) Дж. Цинка, К. Батыгина и Фр. Адамса свела этот
период к сроку из „всего лишь“ 30-100 млрд. лет.[11]
Вычислено, что звезды Млечного пути перейдут на хаотические орбиты только через
1016 (т.е. через десять миллионов миллиардов лет).[12]
Подобной устойчивостью отличаются и другие галактики, даже, может быть, еще самые
первые из них, о которых уже было сказано, что они образовались всего через 400-600
млн. лет после Большого взрыва.
Христианский теизм
Ученые с
натуралистическими убеждениями считают утверждение Бог создал Вселенную весьма упрощенным объяснением вещей, поскольку
для любого сознательного творца было бы очень легко выстроить и организовать
Вселенную. Но так ли это в
действительности?
Повествование в начале Книги Бытия
открывает нам, что Бог сотворил материю
из ничего и развернул Свой грандиозный замысел при построении Вселенной.
Надо, однако, отдавать себе отчет в том, что создание такой динамичной
конструкции – неимоверно сложная задача, поскольку в любой момент она
организована различным образом и при этом сохраняет равновесие. Давайте
вспомним, что только Метагалактика (ее видимая часть) содержит свыше 1022
(десяти тысяч миллиардов миллиардов) звезд и 2000 миллиардов галактик[13],
чье взаимное влияние нужно иметь в виду. (А вся Вселенная, несомненно, гораздо
обширнее.[14])
Равновесие космических систем,
однако, держится на „кончике копья“! В
2009 году ученые сделали более 2 тысяч симуляций движения тел в Солнечной
системе, и во всех этих симуляциях была изменена только орбита самой маленькой
планеты, Меркурия, и притом меньше чем на 1 миллиметр. Оказалось, что в 1% из
случаев даже такое ничтожное изменение делает систему неустойчивой и Меркурий
очень скоро падает на Солнце или сталкивается с Венерой, или же даже
разбалансируется и становится хаотической вся Солнечная система![15]
А в насколько большем проценте случаев система выйдет из строя, если изменить
даже на волосок какую-нибудь из орбит более массивных планет! Это невероятный пример того, с какой исключительной точностью должен был
работать Зодчий Вселенной, чтобы она была устойчивой во времени!
Мы вряд ли отдаем себе отчет, какие
изумительные расчеты должен был проделать космический Зодчий, чтобы привести в
устойчивое динамическое равновесие даже совсем простенькую систему из трех тел.
Прежде всего Он должен был задать такие массы и орбитальные характеристики тел,
чтобы продолжительное время (много миллиардов лет, как мы знаем) их траектории
не стали бы хаотическими и не начали бы пересекаться между собой (т.е. не было
бы столкновений между ними), и в то же время чтобы ни одно из тел не обрело
гиперболическую скорость и не ушло из системы навсегда.
Абстрагируясь от других подробностей,
опишем коротко всю сложность проблемы даже только в отношении гравитации. Для
этой цели процитируем учебник астрономии профессоров Н. Николова и М. Калинкова:
“Самой знаменитой задачей в области небесной механики является задача трех тел, которая была (и остается!) объектом внимания великих математиков и астрономов. Она состоит в
следующем: для некоторого начального момента времени даны координаты и скорости
трех тел с известными массами. Нужно определить положения и скорости этих тел
для произвольного момента времени.
Постановка
задачи для N тел аналогична.
В
действительности расчеты производятся в отношении не тел, а материальных точек. Несмотря на это, достаточно представить, что на каждую материальную
точку действует сложно меняющееся во времени гравитационное поле других точек,
которое в состоянии приводить их к тесным сближениям, чтобы стало ясно, что решение, описывающее невообразимо огромное разнообразие последствий этих сближений, будет иметь чрезвычайно сложный
вид.
Можно
показать, что в задаче для N тел известны только 10 интегралов. Так как для
трех тел существуют 18
дифференциальных уравнений первого порядка, а можно определить только десять интеграционных
констант, задача выглядит неразрешимой. [...]
Общая задача
трех тел была аналитически решена в 1912 году финским ученым Зундманом, который
показал, что изменение координат трех тел, расстояний между ними и времени t
можно представить рядами по степеням
вспомогательной переменной, которые будут абсолютно сходящимися. В 1931-1933
гг. французский ученый Белорицкий установил, что для установления положений
больших планет с точностью астрономических ежегодников нужно из рядов Зундмана
использовать суммы, содержащие не менее 108000 000 членов. Можно считать, что аналитическое решение задачи трех тел найдено,
но оно имеет только теоретическое и ни в коем случае не практическое значение”.[16]
Насколько велико число 108000
000. Если бы во всем объеме наблюдаемой Вселенной не было пустых
пространств и она вся была бы плотно набита элементарными частицами, то, как
вычислили ученые, их число было бы около 10130. Допустим, что на месте
каждой частицы расположен один из членов рядов Зундмана. В таком случае, 10131
будет количество членов в десяти таких вселенных, 10139 – в миллиарде,
10148 – в миллиарде миллиардов, 10157 – в миллиарде миллиардов
миллиардов вселенных и т.д. Таким образом, становится ясно, что существует
огромная бездна(!) до необходимых 108000 000 членов, а при этом
их сумма дала бы нам только приблизительное решение задачи![17]
Ученые заявляют, что с такой задачей не смог бы справиться ни один компьютер,
даже если бы он был размером с Вселенную.
А как можно спроектировать Млечный Путь с его 200-400 миллиардами звезд (плюс звездные скопления, планетные системы и т.д.)? Каждый член галактической “семьи”, если считать его абсолютно твердым
телом (которое не испытывает никаких деформаций), имеет степени свободы, т.е.
может двигаться в трех различных направлениях и вращаться вокруг трех взаимно
перпендикулярных осей. В таком случае, чтобы определить положение тела в
пространстве, нужно знать численные значения трех координат и трех углов
вращения (и следить за скоростью изменения этих параметров во времени). Для
уточнения задачи, однако, нужно заметить, что ни одно из небесных тел не является
абсолютно твердым. Модификации его формы, приливы и отливы меняют скорость его
вращения и направление оси вращения, что воздействует на силы взаимного
притяжения и на орбиты других тел. Нужно учитывать также и: электрические и магнитные взаимодействия; дефект массы (звезды постоянно теряют часть своей массы, которая
превращается в энергию); меняющееся гравитационное поле остальных объектов системы (а даже для трех
тел их координаты и скорости становятся невычислимыми); иногда встречающиеся резонансы (например, между спутниками и планетами в
Солнечной системе); влияние межзвездной среды; некоторые релятивистские эффекты и еще многие другие вещи, которые трудно
даже перечислить.
При поиске
общего решения задачи для совокупности более высоких иерархических образований
(скоплений и сверхскоплений галактик), которые образуют Вселенную, возникает
специфическая трудность, которая заключается в следующем.
Допустим,
что каждая небесная система имеет огромное количество, например N, возможных
упорядоченных состояний при различных величинах масс и орбитальных
характеристик тел, которые содержатся в ней. (Допускаем, что это количество
велико – N, но не бесконечно, поскольку количество вещества и размеры реальных
космических систем ограничены). Если систем две и мы рассматриваем их как
подсистемы одной целостной системы, то в таком случае из-за их взаимных влияний множество упорядоченных состояний всей системы будет сечением только тех
упорядоченных состояний, которые являются общими для обеих подсистем. Если
подсистем три, множество допустимых состояний общей системы еще более
ограничивается – до тех положений, в которых все три подсистемы будут находиться в равновесии. Итак, чем больше подсистем, тем меньшим становится множество их общих равновесных
состояний. Неудивительно, если окажется, что для огромного числа небесных
систем во Вселенной, взятых в целом, может существовать только одна
возможность, при которой все они будут находиться в гармонии между собой и образовывать целостную динамическую структуру Вселенной.
Но в
вышеуказанных рассуждениях мы не учли изменения, которые происходят в каждой
подсистеме. Если система состоит, например,
из двух подсистем, то ее устойчивое состояние не является “механической суммой”
двух устойчивых состояний ее подсистем. (У иерархических структур целое больше
суммы своих частей). Устойчивость каждой из этих подсистем будет уже чем-то
качественно новым, потому что при расчете надо будет учитывать также и внешние
влияния, оказываемые другой подсистемой. В таком случае, новое равновесное
состояние в каждой подсистеме не является подмножеством множества ее
собственных устойчивых состояний (где учтены лишь взаимные влияния между ее собственными телами). Вообще при каждом увеличении
числа подсистем изменяется не только общий порядок всей системы, но и внутренний порядок в каждой подсистеме, потому что все они взаимозависимы и должны
при своем соединении образовывать единую целостную структуру. Если же необходимо спроектировать
Вселенную как единое целое, то при выполнении этого задания нужно следовать заданному плану, в котором все предусмотрено; в противном случае
этот прекрасный “архитектурный храм” обрушится очень быстро.
Все еще
нельзя с уверенностью сказать, какова конструкция Метагалактики,
но следует принять, что она является единой системой, так как все
объекты в ней связаны между собою, будучи интегрированными в сложно
организованные иерархические структуры, которые образуют целостное строение
Вселенной.[18] (фиг. 3).

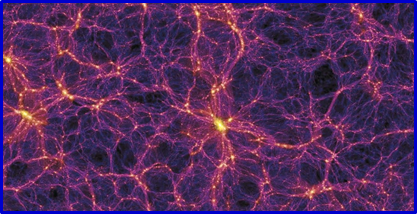
Фиг. 3 а) В рамках программы “Обзор всего неба“ получено изображение более миллиона ярких протяженных объектов, большинство из которых являются галактиками. Вывод, сделанный учеными, состоит в том, что галактики расположены вовсе не хаотически, а организованы в красивые космические системы. б) Часть наблюдаемой крупномасштабной вселенской структуры – скопления и сверхскопления галактик, которые группируются и образуют что-то вроде стенок огромных “клеток”, внутри которых почти отсутствуют подобные образования.[19]
Телескоп “Джеймс Уэбб” позволит нам
в конечном итоге увидеть, что случилось “В начале…”, и
проверить таким образом два варианта, которые мы считаем наиболее вероятными с
точки зрения библейской космологической модели:
А) По велению Божию вещество самоорганизовалось
необыкновенным способом и сформировало хорошо просчитанные планетные, звездные
и галактические системы.
Б) Вселенная была сотворена как
Адам, полностью завершенной и совершенной.
Не будем забывать, однако, что “пути
Господни неисповедимы”, т.е. двери открыты и для сюрпризов!
Если подтвердится библейская
космологическая модель, это значит, что рухнет и вся эволюционная парадигма,
так как она относится и к неорганической материи, как правильно отмечает Дэниел
Деннетт. Таким образом, окончательно будет отброшена и эклектическая теория “теистической эволюции”, которая к
тому же и совершенно еретична с точки зрения христианского вероучения! (См.
гиперссылку, указанную в прим. [20]).
(В этой ситуации, возможно, некоторым
ученым придет в голову вытащить из нафталина так называемую “стационарную модель” Х. Бонди,
Т. Гольда и Ф. Хойла, разработанную еще в 1948 году. В ней вводится специальное
“С-поле”, которое творит вещество так, чтобы
его средняя плотность не изменялась при расширении пространства. Некоторые
сторонники этого взгляда полагают, что мир, возможно, существовал всегда,
причем именно в том хорошо организованном состоянии, в котором мы его наблюдаем
сегодня. Исследования далеких объектов – скоплений, радиогалактик и др. –
опровергли, однако, эту гипотезу, которая не нашла ответ и на вопрос о
происхождении микроволнового фона, а также и на ряд других возражений.
В 1993 году
Ф. Хойл, Дж. Барбидж и Дж. Нарликар создали другую версию, названную ими космологией
квазистационарного состояния, которая тоже ведет к “вечной Вселенной”. Она сумела
объяснить наличие реликтового излучения, его нынешнюю температуру, количество
легких ядер и т.д., но при более внимательном анализе оказалось, что их выводы,
мягко говоря, спекулятивны[21].)
Заключение
Грандиозное построение мироздания побудило Поля Дирака, одного из
крупнейших экспертов по математической физике, воскликнуть: “Можно
было бы описать эту ситуацию, сказав, что Бог – математик высочайшего
класса и что Он использовал довольно сложную математику, чтобы сконструировать
Вселенную. Наши слабые математические познания помогают нам понять лишь малую
часть Вселенной; чем больше будет развиваться математика, тем больше мы можем
надеяться на то, что в будущем будем понимать Вселенную лучше”.[22]
Бог, разумеется, не нуждается в расчетах –
Он обладает совершенным знанием, мудростью и всемогуществом, из-за чего
и вызвал совершенное мироздание к бытию без всякого
интеллектуального и творческого усилия!
Но, как
показывают наблюдения, порядок в этих системах рушится – звезды взрываются,
галактики сталкиваются и т.д. Указанные изменения приводят к резкому изменению во взаимосвязях между членами системы, что в
конечном счете приведет их к гибели. Этот вывод удивительно
хорошо согласуется с библейской точкой
зрения. В Писании сказано, что в начале “тверда Вселенная, не поколеблется” (Пс. 95:10), но из-за грехопадения человека все
творение было подчинено “рабству тления”, т.е. разрушению (Рим. 8: 20, 21).
Если ученые найдут в себе смелость для того,
чтобы переосмыслить данные научной картины мира, то, может быть, они в конце концов признают, что Бог наглядно демонстрирует перед нашими глазами то, как Он построил этот величественный архитектурный ансамбль по имени Вселенная. Недавно
видный британский философ Энтони Флю, которого считали пионером современного атеизма и его защитником целые 50
лет, изменил коренным образом свое мнение по этим вопросам. Говорят, что когда ему задали вопрос, что
случится, если его новый способ мышления будет отвергнут учеными, он ответил: “Мне будет очень жаль. Вся моя жизнь руководствовалась принципом… следовать за доказательствами, куда бы они меня не вели”[23]. Такой принцип мог бы освободить нас
как от наших собственных пристрастий (весьма заблуждающих в большинстве случаев!), так и от обязательной
материалистической парадигмы, из которой проистекает идеологическая
политкорректность в науке.
[1] Вот как толкует первый стих Библии “В начале Бог сотворил небо и землю” один из лучших
знатоков библейского иврита в Болгарии проф. Боян Пиперов: “Творческую деятельность
Бога автор передает еврейским глаголом бара (“сотворил”). Вот что характерно для
употребления этого глагола: 1) он употребляется только для обозначения
божественной деятельности; 2) в результате этой деятельности появляется нечто
чудесное, необыкновенное, новое, небывалое (ср. Исх. 34:10; Иер. 31:22; Пс.
50:12; Ам. 4:13 и другие места, в которых в масоретском тексте стоит повсюду бара).
Этот глагол нигде не употребляется с так называемым в еврейской грамматике
винительным падежом материи (acc. materiae, т.е. “сделать что-то из чего-то”)”.
Пиперов, Б. Тълкуване на
книгата Битие. София: СИ, 1973.
Выражение “из ничего” встречается только в неканонической Второй книге Маккавеев,
где мать ободряет своих сыновей словами: “...Посмотри
на небо и землю и, видя все, что на них, познай, что все сотворил Бог из ничего” (2 Мак. 7:28).
[2]
Dawkins, R. (1996) The Blind Watchmaker. Why the Evidence of the Evolution.
NY: Free Press.
[3] Daniel Dennett, Darwin's Dangerous Idea: Evolution and the
Meanings of Life.1995.
[4] Хокинг, Ст., Л. Млодинов. Великият
дизайн. ИК „БАРД” ООД,
София, 2012, стр. 167-173.
[5] 50 Нобелови лауреати и други велики учени за вярата си в Бога
[6] Еще древние евреи давали себе отчет в том, что Библия не дает
целостного и углубленного описания природного мира. В начале они представляли себе
Землю в качестве плоского диска, накрытого небесным куполом, ниже которого
находится преисподняя (шеол). Позже (IV в. до Р. Х.) еврейские богословы восприняли греческие
космологические представления о сферической Земле, окруженной множеством
концентрических небес. Христиане после II века пользовались геоцентрической системой
Клавдия Птолемея, которая только после XVII века начала постепенно вытесняться гелиоцентрической
моделью Коперника. Это было время, когда христианство рождало в своих недрах
современную науку и в дальнейшем руководствовалось ею, но только в соответствии
с принципом, требующим принимать только эмпирически доказанные теории!
[7] “Vestiges Exist of a Universe Prior to the Big
Bang” – Physicists See a Starkly Different Beginning to the Cosmos
[8]
LISA
[9] Ученые считают хаотическими те системы, которые даже при пренебрежимо
малых событиях могут изменить радикально свое поведение – и таким образом
долгосрочные прогнозы их поведения становятся неосуществимыми. Открытие того,
что параметры хаоса можно измерить, считается третьим большим достижением ХХ века
наряду с теорией относительности и квантовой механикой. Теория хаоса
обладает математическим аппаратом, основанным на поведении некоторых
нелинейных динамических уравнений, чувствительных к начальным условиям. Если
изменить исходные данные даже на ничтожно малые величины, например соизмеримые
с колебаниями числа Авогадро (порядка 10-24), то проверка состояния
системы покажет в результате совершенно разные значения. Примерами таких систем
являются турбулентные потоки в атмосфере, бурное движение воды, биологические
популяции и т.д.
Chaos theory
[10] Drunken solution to the chaotic three-body problem
https://phys.org/news/2021-12-drunken-solution-chaotic-three-body-problem.html
[11] The Great Inequality and the Dynamical Disintegration of the Outer
Solar System
[12] Николов, Н., М. Калинков. „Астрономия”, изд. „Св. Кл. Охридски”,
София, 1998, стр. 242.
[13] The Universe Has 10 Times More Galaxies Than Scientists Thought
[14] Beyond the Universe
https://archive.briankoberlein.com/2016/03/22/beyond-the-universe/
[15] Newton’s three-body problem explained - Fabio Pacucci
https://www.youtube.com/watch?v=D89ngRr4uZg
https://perso.imcce.fr/jacques-laskar/pdf/nature08096&s1.pdf
[16] Николов, Н., М. Калинков. Астрономия.
Изд. „Св. Кл. Охридски”, София, 1998, стр. 76, 77.
[17] Подобным образом можно рассуждать и в отношении атомных ядер. Четыре
взаимодействия (сильное, слабое, электромагнитное и гравитационное), которые
действуют здесь одновременно, создают настолько сложную картину, что невозможно
решить аналитически даже задачу о двух частицах. Но все же давайте предположим,
что в какой-нибудь из бесчисленных вселенных образовалось стабильное атомное
ядро. Если к нему, однако, добавить новый нуклон (протон или нейтрон), оно
немедленно разбалансируется. В таком случае, каждый химический элемент
(соответственно изотоп) должен обладать строго запрограмированным состоянием
ядра для соответствующего количества частиц и переходить к нему автоматически.
[18] Оказывается, что не только в далеком прошлом, когда размеры Вселенной
были достаточно небольшими, чтобы гравитация могла связать воедино все объекты
в ней, но и на гораздо более поздних этапах ее крупномасштабные структуры
остаются связанными каким-то – все еще необъяснимым – способом!
Is the universe controlled by gigantic structures?
https://bigthink.com/hard-science/large-scale-structures/
Если допустить, что гравитационное взаимодействие распространяется
посредством волн (именуемых гравитонами, которые движутся со скоростью света),
то может возникнуть следующее возражение: "Гравитационное излучение
объектов, находящихся в одном конце Вселенной, может быть, еще не дошло до
объектов, расположенных на другом ее конце (из-за ограниченного времени ее
существования), и таким образом не все тела в ней связаны силами
притяжения". На это возражение можно дать следующий ответ: "Каждый объект
во Вселенной взаимодействует хотя бы с теми, которые находятся вокруг него
(как, например, молекулы газа связаны близкодействующими силами), и таким
образом все небесные тела все же объединены гравитационными силами в единую
систему".
Ask Ethan: Do Gravitational Waves Exhibit Wave-Particle Duality?
[19] Народна астрономическа обсерватория “Юрий Гагарин” Стара Загора
[20] СУМЕЛ ЛИ ХОКИНГ СПАСТИ МАТЕРИАЛИЗМ?
[21] Предлагаем читателям главу,
рассматривающую проблемы этих моделей, из “Учебника
космологии”
Эдуарда Райта, переведенного на русский язык:
Ошибки моделей Стационарного состояния и
Квазистационарного состояния
[22] P.A.M. Dirac, "The Evolution of the Physicist's Picture of
Nature," in Scientific American, May 1963, p. 53.
[23] Richard N. Ostling. Famous Atheist Now Believes in God. Associated
Press Newswires, December 9, 2004.